|
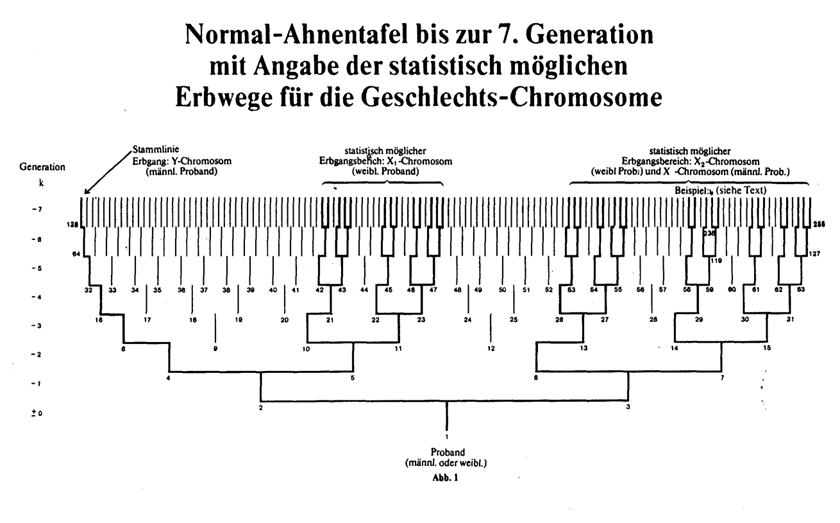
Aufgrund der Gesetzmäßigkeit des X-Y- Chromosomen- Erbganges und der Vorfahren-Struktur zweigeschlechtiger Lebewesen lassen sich in der Aszendenz typische Spaltungs- Proportionen als Erbwahrscheinlichkeitswerte aufstellen. Die Ableitung erfolgt am Beispiel einer menschlichen Normal- Ahnentafel. Ergebnis: Jeder Ahnentafel-Platz ist durch feste "Erbwahrscheinlichkeitswerte" in bezug auf die Geschlechts- Chromosome definiert. Dabei ergeben sich sowohl erbmäßig bevorzugte Ahnentafellinien als auch Ahnentafelbereiche, die für eine Geschlechts- Chromosomenübertragung nicht in Frage kommen können. Zwischen männlichen und weiblichen Ahnentafel- Probanden ergeben sich grundsätzliche Unterschiede. Die Spaltungs-Proportionen folgen in den einzelnen Generationen der sogenannten Fibonacci-Zahlenfolge, und sie stehen damit im engsten Zusammenhang mit dem Verhältnis des sogenannten "Goldenen Schnitts" ( j = 1,618034..), dem in der Natur und der Kunst eine bedeutende Rolle zuzukommen scheint. |
||||||||||||||||||||||||||
|
Über sechs Jahrzehnte sind seit der Entdeckung der geschlechtsgebundenen Vererbung durch Thomas Hunt Morgan (1866-1945) und seinen Mitarbeiter Calvin B. Bridges (1889-1938) an der Taufliege Drosophila melanogaster, dem "Haustier der Vererbungsforscher", vergangen. 1916 konnte Morgan verkünden: "We now know how the factores of heredity carried by the parents are sorted out to the germ cells." Bis heute besteht jedoch in der wissenschaftlichen Genealogie noch immer Unklarheit darüber, ob es erbmäßig bevorzugte Linien auf der Ahnentafel gibt (s. Anm. 1-7). Eher scheint man heute alle Hypothesen und Theorien darüber abzulehnen (s. Anm. 8). Eine Begründung für die erbmäßige Bevorzugung bestimmter Ahnentafellinien soll hier aufgrund der geschlechtschromosomengebundenen Vererbung, im folgenden kurz geschlechtsgebundene genannt, am Beispiel des Menschen gegeben werden. Bekanntlich "mendeln" die Normalchromosome (Autosome) nach der Kernspaltung unabhängig voneinander. Für die Bevorzugung einer oder mehrerer Ahnenlinien gibt es also für sie auch keinen Anhalt. Anders verhält es sich aber bei den Geschlechts-Chromosomen, die eine Sonderstellung einnehmen. Bekanntlich sind sie zunächst Überträger sämtlicher geschlechtsgebundener Erbanlagen. Mag dabei auch das wesentlich größere X-Chromosom die Hauptrolle bei der Anlagen-Übertragung spielen, so ist die "katalytische", vermutlich sogar "dirigistische" Rolle des Y-Chromosoms sicherlich nicht weniger bedeutungsvoll. Die Bedeutung der Geschlechts-Chromosome gegenüber den Autosomen braucht kaum näher beschrieben werden. Es genügt, auf ihre geschlechtsbestimmende Funktion hinzuweisen. Alle primären und sekundären Geschlechtsmerkmale und Geschlechtsunterschiede müssen ursächlich diesen Chromosomen zugeschrieben werden. Inwieweit ein Zusammenspiel mit den Autosomen erfolgt, ist hier für die Ableitung der grundsätzlichen Zahlenverhältnisse von untergeordneter Bedeutung. Da allein die Chromosomen-Kombination XX oder XY darüber entscheidet, ob die Entwicklung zum Mann oder zur Frau erfolgt - von krankhaften Chromosomen-Aberrationen XXY oder XYY soll hier abgesehen werden -, so ist die große Bedeutung dieser Chromosome schon dadurch hervorgehoben. Gleichfalls bedeutsam sind die qualitativen Unterschiede dieser Chromosome innerhalb der Lebewesen einer Gattung. An weitere krankhafte Abweichungen ist dabei zunächst gar nicht gedacht. Bietet doch bereits der "Spielraum des Normalen" eine Fülle faszinierender Möglichkeiten bezüglich individueller Eigenschaftsunterschiede. Die Überträgerrolle der X-Chromosome für gewisse geschlechtsgebundene Krankheiten (z. B. Hämophilie, Rot-Grün-Farbenblindheit, Mikrophthalmie, einige Zahnkrankheiten) ist zur Verfolgung des Erbganges zwar besonders lehrreich (Indikator!) und damit auch für die biologische Genealogie von großem Interesse; außerdem kommt diesem Thema noch große praktische medizinische Bedeutung zu (sehr umfangreiche Literatur!). All dieses kann aber zunächst für die nachfolgenden wahrscheinlichkeitsstatistischen Überlegungen unberücksichtigt bleiben, die Tatsache der Chromosomen-Mutationen und des Crossing over ebenso. Im übrigen zeigen alle Gene im Differentialsegment der Geschlechts-Chromosome nie Crossing over, sie bleiben immer im selben Chromosom und sind Träger der streng geschlechtsgebundenen Vererbung (s. Anm. 9). Der X-Y-Mechanismus gilt sinngemäß für alle zweigeschlechtigen Lebewesen, z. T. mit gewissen Abwandlungen (Bienen). Bei Vögeln und manchen Insekten ist das männliche Geschlecht das gleichanlagige (XX), während bei Fischen und anscheinend allen Säugetieren das weibliche gleichanlagig ist. Der Erbgang der Geschlechts-Chromosome beim Menschen ist bekanntlich wie folgt gekennzeichnet :
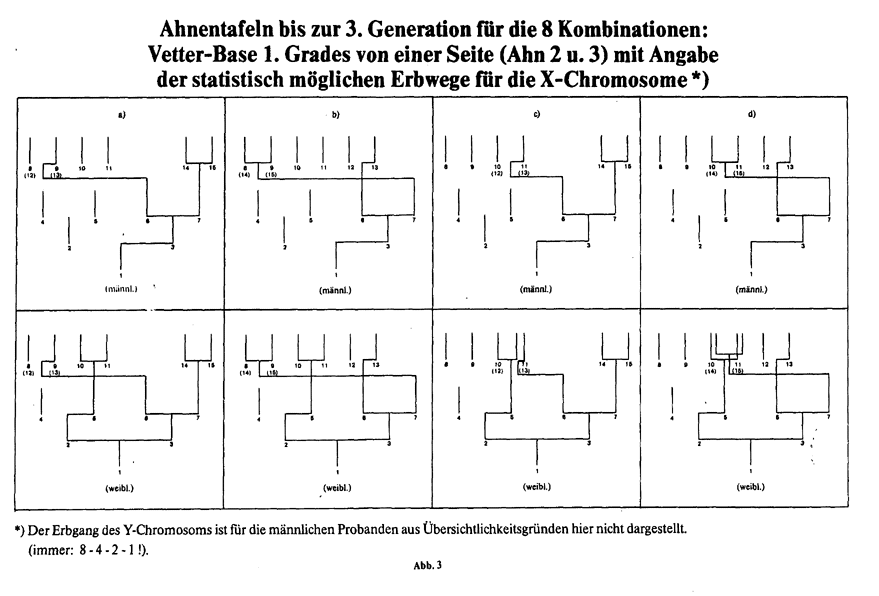
Hieraus ergeben sich nun aber für die einzelnen Filiationslinien unterschiedliche Erbwahrscheinlichkeiten. Dies sei am Beispiel einer menschlichen Ahnentafel erläutert. Wie jede Ahnentafel eines zweigeschlechtigen Lebewesens, besitzt auch die menschliche einen streng gesetzmäßigen Aufbau. Die Nachfahrentafel hat dagegen durch unterschiedliche Kinderanzahl eine unregelmäßige Struktur. Der gesetzmäßige Aufbau der Ahnentafel beruht darauf, daß jedes Individuum nicht mehr und nicht weniger als 2 Eltern besitzt; das ist wohl auch das einzige unumstößliche Gesetz der Genealogie. Die Anzahl der Ahnen in jeder Generation k ist damit eindeutig, nämlich gleich der k-ten Potenz von 2 (2k). Die in Abb. 1 angegebenen Ahnen-Nummern folgen der heute üblichen Nomenklatur nach Kekule, sind aber im übrigen für die hier abzuleitenden Beziehungen ohne Bedeutung; jede andere eindeutige Bezeichnung auf der Ahnentafel führt zum gleichen Resultat. Der "Proband" (männlich oder weiblich) hat hier die Ahnen-Nr. 1, der Vater die Nr. 2, die Mutter Nr. 3, usw. (Abb. 1). Der Vater des Ahns mit Nr. n hat immer 2n als Nr., die Mutter 2n+1. Alle männlichen Personen (außer dem Probanden selbst) haben also immer gerade, alle weiblichen Personen ungerade Ahnen-Nummern. Jede Ahnen-Generation beginnt mit dem Zahlenwert der jeweiligen Potenz von 2 als Ahnen-Nummer. Diese Linie ist im übrigen die Stammlinie, auch reine Manneslinie oder Namenslinie genannt. In Abb. 1 ist das Kind immer mitten unter die beiden Eltern als senkrechter Strich gezeichnet. Soviel zur Struktur und Bezifferung der Ahnentafel. Ein Mann, z. B. hier der Proband Nr. 1 selbst, erhält sein X-Chromosom mit Bestimmtheit (Wahrscheinlichkeit 1) von seiner Mutter (Nr. 3). Sie selbst hat jedoch dieses Chromosom entweder von ihrem Vater (Nr. 6) oder ihrer Mutter (Nr. 7) geerbt, und zwar mit gleicher Wahrscheinlichkeit, so daß die Wahrscheinlichkeitswerte hier in beiden Fällen auf 0,5 abfallen. Bei Ahn Nr. 6 ändert sich dieser Wahrscheinlichkeitswert aber durch Übergang zur nächsten Ahnengeneration nicht, da Ahn Nr. 6 als männliche Person sein X-Chromosom nur von seiner Mutter (Ahn Nr. 13) erhalten kann, während für Ahn Nr. 7 als weibliche Person die Eltern Nr. 14 und Nr. 15 mit gleicher Wahrscheinlichkeit als Überträger infrage kommen können. Die Wahrscheinlichkeitswerte für Ahn Nr. 14 und 15 halbieren sich also wiederum gleichermaßen beim Übergang zur nächsten Ahnengeneration. Das Prinzip der "Disproportionierung" sei damit hinreichend beschrieben. Das Y-Chromosom, das sich immer nur vom Vater auf den Sohn vererbt, hat einen ganz eindeutigen Erbweg auf der Ahnentafel (Stammlinie). Der Wahrscheinlichkeitswert für das Y-Chromosom ist daher innerhalb der Stammlinie generationsunabhängig immer 1. Kann auch bei einem Urenkel z. B. im Einzelfall die ganze Erbmasse des Stammvaters verschwunden sein, bleibt doch sein Y-Chromosom voll erhalten. In der Tabelle (Abb. 2) sind für alle Ahnen bis zur Generation k = -6 die Wahrscheinlichkeitswerte angegeben. Die Nomenklatur der Tabellenwerte ist auch hier der heute in der wissenschaftlichen Genealogie üblichen Form angeglichen. Natürlich sind die in der Tabelle aufgeführten b-Werte für das X-Chromosom (bx für den Mann, bzw. bx1 und bx2 für die Frau) im Einzelfall immer 0 oder 1, [0 oder 1 gilt wegen Crossing over streng genommen nur für die Einzelgene, die auf dem X-Chromosom lokalisiert sind.] und zwar innerhalb der jeweiligen Ahnengeneration nur einmal 1 (bei einem Ahn), dagegen auf allen übrigen Ahnenplätzen 0 (s. Anm. 10). Die mit Strich markierten Ahnentafelplätze scheiden für eine Erbwahrscheinlichkeit der Geschlechts-Chromosome aufgrund des X-Y-Mechanismus von vornherein aus. Es bilden sich "freie Plätze", und zwar in jeder höheren Ahnengeneration "neue" Bereiche. Der prozentuale Anteil der "freien Plätze" wird innerhalb der Ahnenschaft nach rückwärts immer größer. Diese "freien Plätze" können gegebenenfalls bei mehrfacher Verwandtschaft ausgefüllt werden. (z. B. Ahn 12, falls personengleich mit Ahn 10; siehe Abb. 3 c), weibl. Proband). Solche Ahnentafeln, die also Ahnenverlust (auch Ahnenimplex genannt) aufweisen, sollen jedoch bei diesen grundsätzlichen Überlegungen innerhalb der ersten Ahnengenerationen nicht näher betrachtet werden. Freilich, in höheren Ahnengenerationen wird der Ahnenverlust zur Regel, und die "reinen" Zahlenverhältnisse werden wieder überdeckt (z. T. auch durch "Generationsverschiebungen" bedingt, z. B. Nichte-Onkel-Heiraten). In unserem Kulturkreis sind allerdings auch nahe Verwandtenehen durchaus keine Seltenheit. Die 8 verschiedenen Kombinationsmöglichkeiten der Heirat Vetter-Base ersten Grades von einer Seite werden daher in Abb. 3a-d) und Abb. 4 noch dargestellt (statt acht, gibt es hier nur sechs personenverschiedene Urgroßeltern!). In Abb. 4 sind für diese 8 Kombinationen alle Erbwahrscheinlichkeitswerte, beschränkt auf die 3. Generation k = -3, angegeben. Aus Abb. 1 gehen alle möglichen Erbwege für die Geschlechts-Chromosome über 7 Ahnengenerationen anschaulich hervor. Es ist unterschieden nach männlichen und weiblichen Probanden. Für den Erbgang-Einzelfall der mit x1 und x2 bezeichneten X-Chromosome kommt also nur je eine einzige Linie in Frage. Eine solche Linie kann für x2 z. B. sein : 238-119-59-29-14-7-3-1. Außerdem gibt es von der 7. Generation aus weitere 20 mögliche Vererbungswege, z. T. mit größerer, z. T. mit geringerer statistischer Wahrscheinlichkeit. Aus beiden Abbildungen geht hervor, daß zwischen Ahnentafeln mit männlichen und weiblichen Probanden grundsätzliche Unterschiede bestehen. Dem Verfasser ist nicht bekannt, daß auf diese Tatsache in der wissenschaftlichen Genealogie schon einmal näher hingewiesen wurde. Die Ahnentafel eines männlichen Probanden ist gekennzeichnet durch den eindeutigen Erbgang des Y-Chromosoms innerhalb der sogenannten Stammlinie einerseits und den Erbwahrscheinlichkeitsbereich des einzigen X-Chromosoms andererseits. Die Ahnentafel eines weiblichen Probanden ist gekennzeichnet durch zwei unabhängige Erbwahrscheinlichkeits-Bereiche für die beiden X-Chromosome (X1 und X2). Dabei stimmt der X2-Bereich mit dem X-Bereich der "männlichen" Ahnentafel überein. Die möglichen Erbwege der 8 Vetternehen-Kombinationen - hier nur für die X-Chromosomen-Übertragung dargestellt - gehen bis zur 3. Generation aus Abb. 3 hervor. Aus den bX-Werten (mittleren biologischen Verwandtschaftsanteilen) der Tabelle kann man "Spaltungs-Proportionen" p ableiten, die z. B. bei innerhalb der Generation nach der Größe geordneten bx2 -Werten folgende Formen annehmen:
sk ist dabei die Anzahl der bX-Werte in jeder Generation. Vermutlich wurden diese Proportionen durch die vorherrschende Deszendenz-Betrachtungsweise bisher übersehen. Der biometrische und biostatistische Ausbau der Mendelschen Gesetze erfolgte ja meist ziemlich einseitig durch die in die Zukunft gerichtete Frage nach dem "Wohin?". Die Frage nach dem "Woher?" ist aber weniger modern, trotzdem ist sie wohl von gleichem grundsätzlichen wissenschaftlichen Interesse. Auf eine interessante mathematische Zahlenreihe, die in den Wahrscheinlichkeitswerten zutage tritt, sind wir zuletzt gestoßen: Die Anzahlen s der bX-Werte innerhalb der einzelnen Ahnengenerationen folgen nämlich der sogenannten Fibonaccischen Zahlenfolge 1, 1, 2, 3, 5, 8, 13, 21, 34 . . . jedes Glied ergibt sich aus der Summe der beiden vorangehenden: Auch die bx-Erbwahrscheinlichkeitswerte zwischen den weiblichen und männlichen Ahnen stehen in jeder Generation im Verhältnis der Fibonacci-Zahlen. Z. B. in der 7. Ahnengeneration: 13 weibliche und 8 männliche. Geppert (s. Anm. 11) gibt für die Fibonaccische Zahlenfolge nachfolgende Beziehung zu w mit Ableitung (Rückschlußberechnung) an: 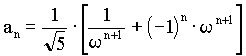 , ,wobei w das "Goldene-Schnitt-Verhältnis" 0,618034.... darstellt. Weiter ist: 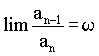 und und 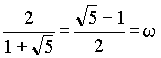 Bei Geppert tritt w bei Auslese- und Inzuchtprozessen in der Deszendenz auf. In der Aszendenz hat meines Wissens erstmals Rösch (s. Anm. 12) auf die Fibonacci-Zahlen hingewiesen, und zwar am extremen Grenzfall einer Inzest-Ahnentafel bei fortgesetzter Eltern-Kinder-Heirat. Die theoretische Ahnenzahl wächst dort in den einzelnen Ahnengenerationen nach dieser Folge. Weiterhin hat Rösch die Fibonacci-Zahlen noch vordergründig als physische Ahnenzahl an der Bienenahnentafel entdeckt (s. Anm. 13). Diese Ahnentafel nimmt aber auch durch ihre Eigengesetzlichkeit eine Sonderstellung ein, da ja dort jedes Bienenmännchen nur von einem Bienenweibchen abstammt, jedes Bienenweibchen aber von einem Bienenelternpaar. Beim X-Chromosomen-Erbgang erscheint diese Zahlenreihe aber auf einer anderen Ebene, es ist die generationsweise Wahrscheinlichkeits-Zahlenfolge in der Normal-Ahnentafel jedes sich gewöhnlich fortpflanzenden zweigeschlechtigen Lebewesens. Rösch (s. Anm. 14) sieht in der Zahl j - dem Kehrwert von w -, übrigens der einzigen positiven Zahl, deren reziproker Wert w sich durch Subtraktion von 1 ergibt 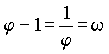 , ,eine ‘Weltkonstante’: "Unsere Zahl j hat solche vielseitige Bedeutung bewiesen, daß sie würdig erscheint, wie die Ludolfsche Zahl p = 3,141592… oder die Basis der natürlichen Logarithmen e = 2,718281… (beide sind ebenfalls unendlich lange Dezimalzahlen ohne Periode) als eine 'Weltkonstante' angesehen zu werden." An Beispielen aus der unbelebten und belebten Natur sowie an Kunstwerken weist Rösch anschaulich auf die Fibonacci-Zahlen und das Goldene-Schnitt-Verhältnis hin. Johannes Kepler nannte die Erkenntnis dieser "Göttlichen Teilung" ein "kostbares Juwel" ! Herr Prof. Dr. W. Meyer zu Capellen, Aachen, wies mich freundlicherweise darauf hin, daß j nicht die einzige positive Zahl ist, die die gleiche Ziffernfolge nach dem Komma hat wie ihr Kehrwert ( es gibt beliebig viele! ). Korrekt ist jedoch: j ist die einzige positive Zahl, deren Kehrwert sich durch Subtraktion von 1 ergibt, was hier im Text von mir bereits korrigiert worden ist. Auf den weiteren theoretischen Ausbau des X-Y-Chromosomen- Erbganges bei Mehrfachverwandtschaft kann hier nicht näher eingegangen werden. Auch die praktische Bedeutung der dargestellten Beziehungen für die Tier- und gegebenenfalls auch Pflanzenzucht soll hier nicht einmal gestreift werden. Dies alles, wie auch eventuelle erkenntnistheoretische Folgerungen, müssen von berufenerer Seite erfolgen. Im Heft 75 sollen die Beziehungen der hier dargestellten Befunde zur Nachfahrenschaft noch kurz behandelt werden. [Der bisher nie veröffentlichte Aufsatz wird hier anschließend im seinerzeit vorgesehenen Wortlaut publiziert.] |
||||||||||||||||||||||||||
|