|
Die erbstatistischen Schlußfolgerungen für den Bereich der Aszendenz haben weitreichende Konsequenzen für die Deszendenz. Diese Problematik soll demnächst in einer umfangreicheren Arbeit dargestellt werden. Einige Ergebnisse werden vorausgenommen und sind hier kurz zusammengestellt. Die Erbstatistik-Strukturen, die für die Vorfahrenschaft abgeleitet wurden (vergl. Archiv für Sippenforschung, C.A. Starke Verlag, Limburg a.d.Lahn (1979) H.74, S.97 f., Abb. 1-2 u. die "Spaltungs-Proportionen") gelten am 2-Kinder-Modell (jeweils 1 Junge und 1 Mädchen) sinngemäß auch für die Nachfahrenschaft. Anstelle des Probanden der Ahnentafel steht jetzt eine Stammperson (Stammvater oder Stammutter); anstelle der Elternpaare stehen Geschwisterpaare. Der Zeitrichtungssinn ist also umgekehrt. Dem 2-Kinder-Modell kommt praktisch und erkenntnistheoretisch eine hohe Bedeutung zu. Das 2-Kinder-Modell enthält alle theoretisch möglichen Filiations-Kombinationen und bietet sich daher als "Meßgerüst" für alle Erbwahrscheinlichkeitswerte an. Natürlich auch für den tatsächlichen Nachfahren-Einzelfall, wo ja jede Filiationslinie fehlen oder auch mehrfach vertreten sein kann. Als Nachfahren-Zusatzbezeichnung ist ein Code zweckmäßig, aus dem die Filiationslinien-Kombination Mann-Frau-… in Bezug auf die Stammperson bündig hervorgeht, ähnlich der Kekule-Nr. für die Vorfahrenschaft. Aus den Filiationslinien-Kombinationen Mann-Frau-... ergeben sich bei einfacher Verwandtschaft alle bx-Werte allgemein als 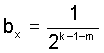 k ist die jeweilige Ahnen- bzw. Nachfahrengeneration; m ist die Anzahl der männlichen Personen, wobei das Anfangsglied (Proband bzw. Stammperson) und das Endglied nicht mitzuzählen sind. Bei zwei oder mehr aufeinanderfolgenden männlichen Personen wird bx immer zu 0. Bei Mehrfachverwandtschaft ergeben sich die bx-Werte als Summe der Einzel-bx-Werte aus allen Filiationslinien- Kombinationen (Erbwegen). Auch für die Nachfahrenschaft wird der prozentuale Anteil derjenigen Personen, die für eine X-Chromosomen-Übertragung von der Stammperson niemals in Frage kommen können (aufgrund ihrer besonderen Filiationslinien-Kombination) von Generation zu Generation gegenüber der Gesamtpersonenzahl immer größer ("freie Plätze") . In einer idealen konstanten stationären Bevölkerung gilt die wichtige Beziehung : "Wieviel Vorfahren, soviel Nachfahren" (Praetorius1 und v.Schelling2). Dieser Beziehung liegt auch wieder das 2-Kinder-Modell zugrunde. Werden die Personenzahlen ohne Ahnen- oder Nachfahrenschwund (Implex) betrachtet, dann wächst die Zahl der Nachfahren im statistischen Mittel nach dem "Gesetz der großen Zahlen" ebenso mit der Generations-Nr. , wie nach rückwärts die Zahl der Ahnen, nämlich nach Potenzen von 2. Diese Beziehung wird aber bei einer Nachfahrenschaft nur von denjenigen Personen erfüllt, die bis zum Betrachtungszeitraum (z.B. Gegenwart) selbst noch Nachfahren besitzen. Diese Nachfahrenpersonenzahl, die sich von der sog. "wirksamen" Nachfahrenzahl nw -definiert als "alle Nachfahren, die Kinder haben" (Praetorius 1911 und 1959)- grundsätzlich unterscheidet, sei hier zunächst als "lebendige" Nachfahrenzahl nl(k) bezeichnet. Die "lebendige" Nachfahrenzahl nl( k) umfaßt also nur solche Personen, die durch alle Generationen bis zur jüngst betrachteten Zeit (meist Gegenwart) noch Nachkommen haben. Es gilt, wenn wir unter a(k) die theoretische Ahnenanzahl verstehen: Der Begriff der "lebendigen" Nachfahren ist aber nicht nur theoretisch von Interesse, er hat auch eine hohe genetische Bedeutung. Denn das Erbgut jeder Bevölkerung ist ausschließlich von "lebendigen" Nachfahren gebildet worden. Jede Ahnentafel eines z.B. heute lebenden Probanden kann natürlich nur aus solchen "lebendigen" Nachfahren bestehen. Oder kurz: "Alle Ahnen sind 'lebendige' Nachfahren!". Verschiedene neue genetische und genealogische Aufgabenstellungen werden sich aus der Erbstatistik der Geschlechts-Chromosomen ergeben. An der abgeleiteten Erbstatistik-Struktur (b-, bx- und by-Werte) läßt sich das "Grundgesetz" der Populationsgenetik, das sog. Hardy-Weinberg-Gleichgewicht sehr leicht ableiten. Im 2-Kinder-Modell führt für jedes Chromosom der Stammperson (sowohl Normalchromosom als auch Geschlechtschromosom) nämlich statistisch nur eine einzige Erblinie in die Nachfahrenschaft. Dadurch bleibt der relative Chromosomen-Anteil bei konstanter Bevölkerung gleich. Auch bei einer abnehmenden oder zunehmenden Bevölkerung ändert sich am relativen Anteil der Chromosomen bzw. Gene nichts da die Erblinien statistisch im gleichen Verhältnis abnehmen oder zunehmen wie die Gesamtnachkommenschaft. Daß auch der relative Anteil der Chromosomen-Kombinationen bzw. der einzelnen sich vertretenden Erbfaktoren (Allele) gleich bleiben muß, versteht sich eigentlich fast von selbst. Abstrahieren wir und denken bei wahlloser Vermischung (wissenschaftlich: Panmixie oder random mating) an gleich große weiße und schwarze Kügelchen anstelle der Chromosomen bzw. Erbfaktoren in möglichst großer Zahl. Eine gegenseitige Berührung von Kügelchen soll für die resultierenden Erbkombinationen stehen. Das statistische Verhältnis der Kügelchen-Berührungen (Punkte): wird in einem gut geschüttelten Becher jedoch immer dasselbe sein, sofern nur das relative Verhältnis der schwarzen zu den weißen Kügelchen immer dasselbe bleibt. Das ist das ganze, doch ziemlich einfache Prinzip des Hardy-Weinberg-Gleichgewichtes. Unabhängig von der ursprünglichen relativen
Verteilung eines spezifischen X-Chromosomenanteils bzw. der dort
lokalisierten Erbfaktoren in einer Bevölkerung (z.B. rassespezifische
Gene oder mutierte Gene) -also gleichgültig, ob ursprünglich
nur auf männliche, weibliche oder ein beliebiges Verhältnis
männlich : weiblich verteilt- strebt die X-Chromosomen-Verteilung
einem Grenzwert 1:2 zu, d.h. Bei einer ungleich auf die Geschlechter verteilten Chromosomenverteilung, also den X-Chromosomen (1:2) und den Y-Chromosomen (1:0), muß eine wirksame Abweichung vom Geschlechterverhältnis 1:1 daher einen Evolutionsfaktor hinsichtlich dieser Chromosomen darstellen! Literatur
geschrieben: München, 7. August 1979 |